引言
鞭毛是纤毛或鞭毛上的毛发状侧翼附属物,参与机械感觉和细胞运动,但其组成和结构尚不清楚。
2024年3月28日,清华大学/深圳医学科学院颜宁、清华大学闫创业及潘俊敏共同通讯在
Cell 在线发表题为“
Structure-guided discovery of protein and glycan components in native mastigonemes”的研究论文,该研究报道了从莱茵衣藻纤毛中分离的天然纤绒毛 (Mastigoneme) 3.0 Å分辨率的冷冻电镜结构。
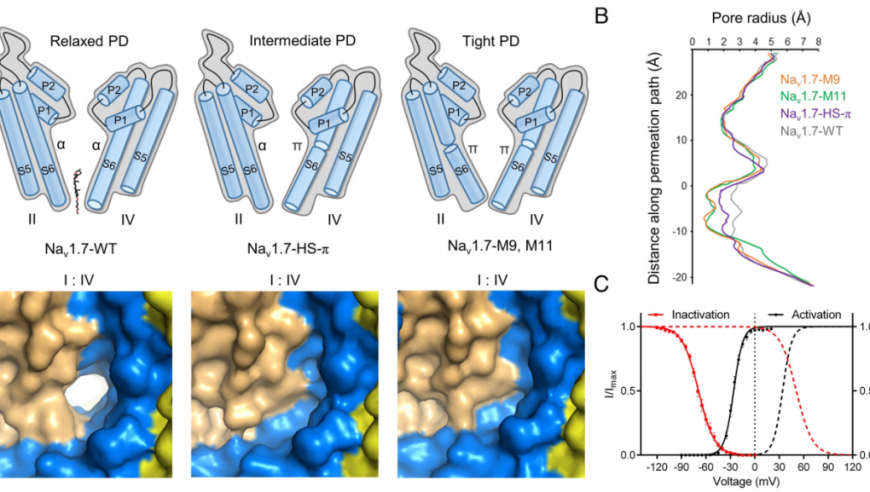
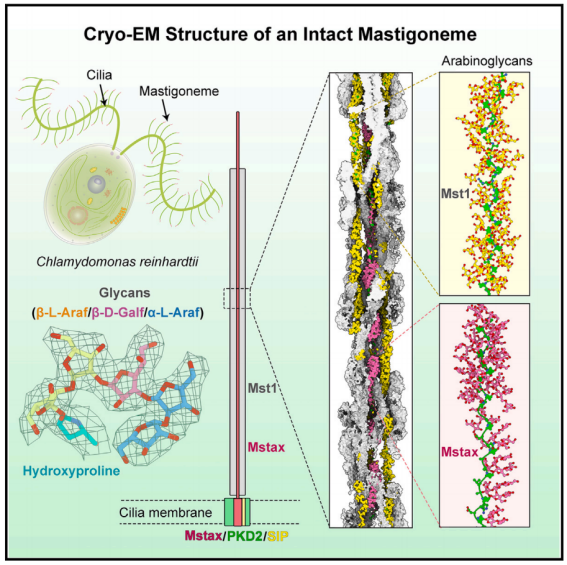
长茎形成一个超级螺旋,每个螺旋旋转由四对反平行的纤绒毛素样蛋白1 (Mst1)组成。在Mst1中,在II型聚羟基脯氨酸(Hyp)螺旋周围发现了大量阿拉伯糖聚糖,它们代表了植物和藻类中常见的一类糖基化。EM图谱揭示了纤绒毛轴心蛋白(Mstax),该蛋白富含重度糖基化的Hyp,并含有PKD2样跨膜结构域(TMD)。Mstax有近8000个残基,从细胞内区域到纤绒毛素的远端,为Mst1的组装提供了框架。该研究提供了对天然生物结构中蛋白质和聚糖相互作用复杂性的见解。
纤毛,或鞭毛,突出到细胞外环境服务于双重目的检测环境的变化和促进细胞运动。
一些单细胞原生生物的纤毛排列有侧毛,称为纤绒毛。先前的研究表明,纤绒毛可能参与机械刺激的感知。
它们还可以通过增加纤毛在蛙泳运动时的有效表面积来增强纤毛推进力。
有管状和非管状纤绒毛。
管状纤绒毛属通常长1毫米,直径15-19纳米,相对坚硬,而非管状纤绒毛属较短(0.8毫米),较薄(10纳米),柔韧,易脱落。衣单胞菌纤毛上的非管状纤绒毛素被认为与多囊肾病蛋白(PKD2)(也称为多囊素-2 [PC-2])有关,多囊素-2是一种瞬时受体电位(TRP)家族通道,其跨膜区域共享电压门控离子通道(VGIC)折叠。在哺乳动物细胞的初级纤毛上,PKD2的三个亚基与PKD1(也称为PC-1),(具有巨大的多结构域胞外区域)组合形成异源四聚体。
然而,PKD1/PKD2异构体的生理功能尚不完全清楚,该异构体是否具有通道活性仍存在争议。
预测衣藻上的纤绒毛是由重度糖基化的纤绒毛样蛋白1 (Mst1)组成的螺旋状聚合物。
由于纤绒毛的高分辨率结构的缺乏,完整的组件,准确的组装,以及与PKD2潜在相互作用的细节仍然是未知的。
此外,聚糖的结构或功能作用仍然难以捉摸。
糖基化是翻译后修饰(PTM)的一种主要形式,它显著地扩展了蛋白质的结构复杂性和功能多样性。
糖化蛋白参与多种生物过程,如信号传导、免疫反应和细胞分化。在核酸、蛋白质、脂质和碳水化合物这四种主要的生物分子中,后两者是结构生物学最具挑战性的目标。糖基化通常通过点突变或去糖基化来去除,以改善基因的均匀性蛋白质结晶。使用单粒子冷冻电镜(cryo-EM)进行结构分析,可以保存糖基化。
然而,由于多糖链固有的柔韧性,在EM图中只能分辨出靠近修饰残基的几个糖基。碳水化合物结构信息的缺乏严重限制了人们对一类主要生物分子的理解。
模式图(Credit:
Cell)
为了扩大人们对糖蛋白和天然生物结构的了解,利用单粒子冷冻电镜(cryo-EM)研究了莱茵衣藻(
Chlamydomonas reinhardtii)内源纤绒毛的组装,并获得了3.0 Å的重建。
除了交织在一起的Mst1原纤维的高阶低聚物外,出色的EM图谱还能够鉴定出作为纤绒毛中心轴的蛋白质。研究人员将这种高度糖基化、富含羟脯氨酸(Hyp)的大蛋白命名为纤绒毛特异性轴蛋白Mstax。对Mstax未解析片段的结构预测显示,在近膜区和跨膜区存在PKD2样褶皱,从而为纤绒毛和PKD2之间的相互作用提供了合理的分子基础。
该研究提供了对天然生物结构中蛋白质和聚糖相互作用复杂性的见解。
https://www.cell.com/cell/abstract/S0092-8674(24)00240-X
|探索君
排版|探索君
文章来源|“iNature”